
Journal
of
Physiology
(1996),
492.3,
pp.689-703
Calcium
handling
and
purinoceptor
subtypes
involved
in
ATP-induced
contraction
in
rat
small
mesenteric
arteries
G.
J.
L.
Lagaud,
J.
C.
Stoclet
and
R.
Andriantsitohaina
*
Laboratoire
de
Pharmacologie
et
Physiopathologie
Cellulaires,
Universite
Louis
Pasteur
de
Strasbourg,
URA
CNRS
600
Faculte
de
Pharmacie,
BP
24,
67401
Illkirch-Cedex,
France
1.
The
relationship
between
the
stimulation
of
ATP
receptors,
the
increase
in
intracellular
free
calcium
concentration
([Ca2+]1;
measured
using
the
fluorescent
indicator
fura-2),
contraction
and
the
subtypes
of
purinoceptors
involved
were
investigated
in
the
small
mesenteric
artery
of
the
rat.
2.
In
normal
physiological
solution,
ATP
(0
001-3
mM)
caused
concentration-dependent
increases
in
both
[Ca2+]i
and
contraction.
Both
responses
produced
by
ATP
(1
mm)
were
inhibited
by
50
%
in
the
presence
of
nitrendipine
(1
/M)
and
were
abolished
in
the
presence
of
nitrendipine
plus
SK&F
96365
(30
/sM).
3.
In
Ca2+-free
medium,
ATP
(3
mM)
elicited
a
transient
increase
in
both
[Ca2+]i
and
tension
which
were
abolished
by
caffeine
and
decreased
by
65%
by
thapsigargin
(1
/SM).
Moreover,
ATP
(1
and
3
mm)
produced
increases
in
the
[3H]D-myo-inositol
1,4,5-trisphosphate
([3H]IP3)
content
of
vessels
in
a
concentration-dependent
manner.
4.
Treatment
of
the
vessels
with
Bordetella
pertussis
toxin
(PTX)
inhibited
contractions
to
ATP
linked
to
the
influx
of
calcium
through
nitrendipine-sensitive
mechanisms,
but
not
those
linked
to
the
release
of
Ca2+
from
intracellular
stores
nor
the
capacity
of
ATP
in
increasing
IP3
content
of
the
vessels.
5.
The
order
of
potency
of
ATP
and
its
analogues
in
eliciting
contraction
was
a,,fl-methylene-
ATP
(a,/3-MeATP)
>
2-methylthio-ATP
(2-MeSATP)
>
ATP
=
ADP.
The
response
to
ATP
was
inhibited
by
suramin.
Reactive
Blue
2
(up
to
100
uM)
did
not
affect
the
contractile
response
to
ATP.
Pyridoxal-phosphate-6-azophenyl-2',4'-disulphonic
acid
4-sodium
(PPADS)
and
a,,i-MeATP
abolished
the
response
to
low
concentrations
of
ATP
and
reduced
contractions
elicited
by
high
concentrations
of
ATP.
6.
After
blockade
of
P2x-purinoceptors
with
PPADS,
the
order
of
potency
of
ATP
and
its
analogues
was
2-MeSATP
>
ATP
=
ADP.
UTP
produced
concentration-dependent
contractions
which
were
not
affected
by
suramin,
Reactive
Blue
2,
PPADS
or
a,fl-MeATP,
suggesting
the
presence
of
P2U-purinoceptors.
7.
The
results
suggest
that
low
concentrations
of
ATP
activate
P2x-purinoceptors
and
produce
an
influx
of
calcium
through
both
voltage-dependent
calcium
channels
sensitive
to
nitrendipine
and
through
receptor-operated
calcium
channels
sensitive
to
SK&F
96365.
High
concentrations
of
ATP
activate
P2Y-purinoceptors
which
promote
firstly
a
nitrendipine-sensitive
calcium
influx
via
a
PTX-sensitive
G
protein
and
secondly
a
release
of
Ca2P
from
an
internal
source
via
the
production
of
1P3.
ATP
is
an
extracellular
messenger
which
acts
as
a
neuro-
nervous
system
of
different
blood
vessels,
ATP
is
involved
transmitter
or
co-transmitter
in
the
central
and
peripheral
in
the
non-adrenergic,
non-cholinergic
component
of
the
nervous
systems
(Von
Kiiglegen
&
Starke,
1991;
Barnard,
nerve-evoked
vasoconstriction
(Hoyle
&
Burnstock,
1991).
Burnstock
&
Webb,
1994).
In
the
perivascular
sympathetic
The
mechanism
by
which
ATP
produces
contraction
is
*
To
whom
correspondence
should
be
addressed.
4639
689

G.
J
L.
Lagaud
and
others
complex
and
varies
with
species,
tissue
and
experimental
model.
The
contraction
induced
by
ATP
is
generally
associated
with
depolarization
(Benham,
1989;
Juul,
Plesner
&
Aalkjaer,
1992)
which
can
be
due
to
direct
opening
of
cationic
non-selective
channels,
or
to
opening
of
voltage-dependent
calcium
channels,
or
both
(Von
der
Weid,
Serebryakov,
Orallo,
Bergman,
Snetkov
&
Takeda,
1993).
In
some
cases
ATP
can
induce
the
release
of
Ca2P
from
the
inositol
trisphosphate-sensitive
stores
(Tawada,
Furukawa
&
Shigekawa,
1987).
Both
these
effects
of
ATP
riesult
in
an
increase
in
intracellular
free
calcium
concentration
([Ca2+]1)
which
leads
to
activation
of
contractile
proteins.
Furthermore,
G
proteins
might
be
involved
in
the
contraction
induced
by
ATP
(McMillan,
Soltoff,
Cantley
&
Talamo,
1987;
Dubyak,
Cowen
&
Meuller,
1988;
Ohya
&
Sperelakis,
1989).
ATP
mediates
its
effects
through
activation
of
P2
-
purinoceptors
that
have
been
divided
into
two
subtypes,
P2Xand
P2Y
This
classification
is
based
on
pharmacological
data
which
take
into
account
the
rank
order
of
agonist
potency
using
structural
analogues
of
ATP
(being
a,
,
-
methylene
-
ATP
(a,/3
-
MeATP)
>
2
-
methylthio
-
ATP
(2-MeSATP)
>
ATP
=
ADP
for
P2x-purinoceptors)
and
the
activity
of
antagonists
(Burnstock
&
Kennedy,
1985).
Recently,
other
P2-purinoceptor
subtypes
have
been
reported,
such
as
P2T
on
platelets,
P2U
on
smooth
muscle
cells
and
P2Z
on
mast
cells
(for
review
see
Fredholm
et
al.
1994;
Barnard,
Burnstock
&
Webb,
1994).
However,
the
use
of
ectonucleotidase
inhibitors,
enzymes
which
hydrolyse
ATP
to
adenosine,
and
also
data
from
cloning
studies
bring
some
controversy
in
the
classification
of
P2-
purinoceptors.
Indeed,
inhibitors
of
ectonucleotidases
such
as
FPL
67156
(6-N,N,-diethyl-D-/3,y-dibromomethylene-
ATP),
as
well
as
suramin
and
FPL
66301
(/3,y-methylene-
dibromo-2-methylthio-L-ATP),
influence
greatly
the
rank
order
of
agonist
potency
at
P2X-purinoceptors
in
rabbit
ear
artery
and
in
guinea-pig
vas
deferens
(McKechnie,
Crack,
Dainty
&
Leff,
1994;
Sneddon,
Walker,
Leff
&
Kennedy,
1994;
Crack,
Beukers,
McKechnie,
IJzermann
&
Leff,
1994).
In
these
preparations,
ATP
and
2-MeSATP
become
more
potent
than
a,,-MeATP
in
eliciting
contraction.
Mloreover,
the
order
of
agonist
potency
of
the
newly
cloned
P2X-purinoceptors
from
rat
vas
deferens
(2-MeSATP
>
ATP
>
a,fl-MeATP
>>
ADP;
Valera
et
al.
1994)
and
from
phaeochromocytoma
or
PC12
cells
(ATP
>
ATPyS
>
2-
MeSATP;
Brake,
WTagenbach
&
Julius,
1994)
differ
from
that
previously
described
by
Burnstock
&
Kennedy
(1985).
Recently,
Burnstock
and
his
co-workers
(Fredholm
et
al.
1994;
Barnard
et
al.
1994)
proposed
a
new
classification
system
based
on
the
structure
of
the
receptor
which
is
either
a
ligand-gated
ion
channel
designated
as
P2X
or
a
G
protein-coupled
receptor
designated
as
P2Y.
Thus,
the
classification
of
purinoceptors
has
become
complex
and
needs
to
be
reassessed,
taking
into
account
a
combination
of
both
molecular
structure
and
subtype
selective
drug
action.
In
general,
studies
concerning
the
effects
of
ATP
have
been
performed
on
vascular
smooth
muscle
cells
in
culture
or
on
large
vessels.
Few
studies
have
been
performed
on
resistance
arteries
which
are
involved
in
the
regulation
of
blood
pressure.
In
small
mesenteric
resistance
arteries
of
the
rat
using
different
analogues
of
ATP,
ATP
has
been
shown
to
mediate
contraction
via
activation
of
P2X-
purinoceptors.
This
contraction
is
associated
with
depolarization
and
a
subsequent
rise
in
intracellular
[Ca2+]i
(Juul
et
al.
1992).
However,
the
mechanism
by
which
ATP
produces
the
rise
in
[Ca2+]i
in
this
preparation
is
not
fully
understood
nor,
because
of
the
complexity
of
the
classification
of
purinoceptors,
has
the
exact
identity
of
the
P2-purinoceptor
subtypes
involved
been
elucidated.
Therefore,
the
aim
of
the
present
study
was
to
further
investigate
P2-purinoceptor
subtypes
activated
by
ATP
and
their
coupling
mechanims
with
[Ca2+]i
and
the
resultant
contraction
in
small
mesenteric
artery
of
the
rat.
METHODS
Arterial
preparation
and
mounting
Alale
WVistar
rats
(250-350
g;
bred
in
the
authors'
institute)
were
killed
by
cervical
dislocation
and
exsanguinated
by
carotid
artery
transection.
The
viscera
were
exposed,
and
a
proximal
segment
of
the
small
bowel
was
removed
and
pinned
in
a
dissecting
dish
containing
physiological
salt
solution
(PSS)
of
the
following
composition
(mM):
NaCl,
119;
KCl,
4
7;
KH2P0
4;
NaHCO3,
14-9;
MgSO4,
1-17;
CaCl2,
2-5;
glucose,
5-5.
Branch
II
oI
III
mesenteric
resistance
arteries
were
cleaned
of
fat
and
connective
tissue,
and
a
segment
2
mm
long
was
removed.
The
segment
was
then
mounted
in
a
myograph
filled
with
PSS
kept
at
37
0C
and
continuously
gassed
with
a
mixture
of
95%
02-5%
CO2
(pH
7
4)
(Andriantsitohaina,
Andre
&
Stoclet,
1990).
Briefly,
two
tungsten
wires
(30
/um
diameter)
were
inserted
through
the
lumen.
Miechanical
activity
was
recorded
isometrically
by
a
force
transducer
(Kistler-Morse,
DSG
BE4;
Kulite
Inc.,
NJ,
USA)
connected
to
one
of
the
two
tungsten
wires,
the
other
being
attached
to
a
support
carried
by
a
micromanipulator.
In
some
experiments
small
resistance
arteries
were
incubated
for
12
h
at
37
0C
in
mimimun
essential
medium
(AIEM)
containing
10%
fetal
calf
serum
and
600
ng
mil-
Bordetella
pertussis
toxin
(PTX)
that
wvas
gassed
with
5%
C02-95%
02-
In
control
experiments
vessels
were
incubated
with
PTX-free
medium
under
the
same
conditions.
Once
mounted,
the
vessel
was
equilibrated
for
30
min
before
being
passively
stretched
to
an
internal
diameter
that
yielded
a
circumference
equivalent
to
90%
of
that
given
by
an
internal
pressure
of
100
mmnHg;
this
required
a
load
of
about
200
mg.
The
internal
diameter
of
the
vessels
used
in
this
study
ranged
between
150-200
/um.
After
setting
the
v-essel
to
its
working
length,
it
was
challenged
twice
with
10
/LM
noradrenaline
to
elicit
reproducible
contractile
responses.
The
presence
of
functional
endothelium
was
assessed
in
all
preparations
by
the
ability
of
acetylcholine
(1
,UM)
to
induce
more
than
50%
relaxation
of
vessels
pre-contracted
with
noradrenaline
(10
/tM).
Unless
otherwise
indicated
the
experiments
were
performed
on
vessels
with
intact
endothelium.
J
Physiol.
492.3
690

Ca2+
and
A
TP
in
resistance
arteries
Contraction
experiments
Effect
of
ATP
in
normal
PSS
and
in
Ca2+-free
medium
on
control
and
PTX-treated
vessels.
Concentration-response
curves
to
ATP
(0
001-3
inm)
vere
constructed
by
non-cumulative
application
in
either
normal
PSS
or
in
Ca2+-free
medium.
For
Ca2+-
free
PSS,
calcium
was
omitted
and
0
5
mM
EGTA
was
added.
Additions
of
different
concentrations
of
ATP
were
separated
by
45
min
washout
periods.
These
experiments
showed
that,
in
Ca2+-
free
PSS,
ATP
was
able
to
produce
contraction
only
at
a
concentration
of
3
mni.
Thus,
in
order
to
distinguish
extracellular
and
intracellular
Ca2P
components
of
the
ATP-induced
contraction,
ATP
was
used
at
a
concentration
of
either
1
or
3
mai
in
the
following
experiments.
In
order
to
study
the
effect
of
ATP
metabolites
such
as
ADP,
AMIP
and
adenosine,
concentration-response
curves
to
ADP
(1-3000
/LM),
AMP
(1-3000
AM)
and
adenosine
(1-3000
tmI)
were
constructed
by
non-cumulative
application
in
normal
PSS.
Also,
the
relaxant
effect
of
AMP
and
ADP
was
investigated
on
vessels
that
had
been
pre-contracted
with
3
/uM
noradrenaline.
Calcium
entry
blockers
were
used
in
order
to
study
the
Ca2P
entry
component
of
the
ATP-induced
contractions.
They
were
applied
at
maximally
active
concentrations,
being
1
JUM
for
the
voltage-
operated
calcium
channel
blocker,
nitrendipine
and
30
/M
for
the
receptor-mediated
Ca2P
entry
blocker,
SK&F
96365
(1-{/-[3-
(4-
methoxyphenyl
)
propoxy]-
4
-methoxyphenethyl
-1H-iinidazole
hydrochloride)
(AMerrit
et
al.
1990).
These
compounds
wNere
used
under
two
different
experimental
conditions.
In
the
first
experimnental
conditioin,
their
effects
were
studied
either
on
the
contractile
response
to
ATP
(1
mM)
in
normal
PSS
or
on
the
contractile
responses
of
vessels
exposed
to
1
mm
ATP
in
Ca2P-free
medium.
In
the
second
experimental
condition,
the
experiments
were
performed
after
depletion
of
intracellular
Ca2P
stores
by
repeated
exposure
of
the
vessels
to
noradrenaline
(10
/AM),
in
a
Ca2+-free
PSS
containing
2
mM
EGTA,
until
there
was
no
contraction
to
noradrenaline.
Ca2+
entry
blockers
were
pre-
incubated
with
the
vessel
for
30
min
before
addition
of
ATP.
A
washout
period
of
45
min
was
allowed
between
each
experimental
protocol.
To
study
the
component
of
the
ATP-induced
contractions
due
to
internal
Ca2+
release,
two
intracellular
Ca2+
mobilizing
agents
were
used
at
maximally
active
concentrations
on
rat
mesenteric
resistance
arteries
as
described
previously
(Shima
&
Blaustein,
1992;
Andriantsitohaina,
Lagaud,
Andre,
Muller
&
Stoclet,
1995);
these
agents
were
thapsigargin
(1
AM),
an
inhibitor
of
calcium
ATI'lase,
and
caffeine
(10
m),
an
activator
of
the
Ca2+-induced
Ca2+
release
channels.
The
experiments
were
performed
in
Ca2+-
free
medium.
The
intracellular
Ca2P
stores
were
loaded
with
Ca2+,
by
stimulating
the
vessels
with
a
depolarizing
solution
i.e.
100
mx
KCl-PSS
containing
2
5
mM
CaCl2
in
which
KCl
was
substituted
for
an
equimolar
amount
of
NaCl.
ATP
was
used
at
a
concentration
of
3
mx.
Thapsigargin
was
pre-incubated
for
30
min
before
addition
of
ATP.
Effect
of
different
purinoceptor
agonists
and
antagonists.
Concentration-effect
curves
for
different
purinergic
analogues
(a,fl-MeATP,
ADP,
2-MeSATP,
uridine
5'-triphosphate
(UTP))
were
constructed
by
non-cumulative
application
of
the
drugs
using
serially
increasing
concentrations.
Each
concentration
of
agonist
w^as
applied
for
a
2-3
inin
period
during
which
a
peak
effect
was
reached.
All
agonists
were
added
allowing
a
45
min
washout
The
effects
of
different
purinoceptor
antagonists
were
examined
on
concentration-response
curves
to
ATP
and
UTP.
Inhibitors
used
Xwere
the
non-selective
P2-purinoceptor
antagonist
suramin
(1-100
AM),
the
P2Y-selective
antagonist
Reactive
Blue
2
(1-
100
#uM),
the
newly
introduced
selective
P2X-antagonist
pyridoxal-
phosphate-6-azophenyl-2',4'-disulphonic
acid
4-sodium
(PPADS,
1
-100
tM)
and
the
P2X-desensitizing
agent
a,fl-MeATP
(10
/tM).
To
study
the
non-P2x-component
responses,
concentration-effect
cuives
for
ATP,
ADP
and
2-MeSATP
were
constructed
in
the
presence
of
maximally
active
concentrations
of
PPADS
(60
ymzi).
Also,
responses
to
maximally
active
concentrations
of
the
selective
P2Y-agonist
adenosine-5'-O-(2-thiodiphosphate)
(ADP/S,
300
/M)
were
elicited
in
normal
PSS
in
the
absence
and
presence
of
PPADS
(60
/vvi)
or
Reactive
Blue
2
(100
uM)
and
in
CPa2-ftee
PSS.
All
the
inhibitors
were
pre-incubated
at
37
°C
foi
30
min
before
the
addition
of
ATP
and
they
were
present
in
the
bath
solution
throughout
the
experiments.
Intervals
of
45
min,
with
a
wash
every
10
min,
were
allowed
between
consecutive
application
of
inhibitors.
Measurement
of
[Ca2+]i
Simultaneous
measurements
of
[C'a2]i
and
contraction
were
performed
in
order
to
determine
directly
the
relationship
between
the
stimulation
of
ATP,
the
increase
in
[Ca2+]i
and
the
contraction
of
the
small
mesenteric
arteries.
Changes
in
[Ca2+]i
were
determined
by
measuring
the
fluorescence
of
trapped
fura-2
with
a
dual
excitation
wavelength
fluorometem
(Fluorolog
II;
SPEN,
Edison,
NJ,
USA)
using
the
method
described
by
Bukoski,
Bergmann,
Gairard
&
Stoclet
(1989).
The
vessel
segments
were
loaded
with
fura-2
by
incubation
in
the
(lark
in
PSS
containiing
5
/LM
fura-2
AM
(the
acetoxymethyl
ester
of
fura-2)
and
20%
pluronic
acid
for
2
h.
PSS
kept
at
37
°C
and
continuously
gassed
with
95%
02-5%
CO2
mixture
(pH
7
4)
was
used
in
this
study
instead
of
Hepes-buffered
salt
solution
used
by
Bukoski
et
al.
(1989).
At
the
end
of
each
experimilent,
the
Ca2+
signal
was
calibrated
using
ionomycin
(20
jtU),
noradrenaline
(101tAi)
anld
Ca2+
(5
mM)
for
the
maximal
fluorescence
and
20
mM
EG'T'A
in
Ca2+-free
solution
for
the
minimal
fluorescence.
The
ratio
of
fluorescence
of
the
emission
of
fura-2
obtained
at
510
nm
was
calculated
after
subtraction
of
the
autofluorescence
at
340
and
380
nm.
Measurement
of
inositol
1
,4,5-trisphosphate
(P3)
content
Tritiated
1P3
was
determined
using
thie
method
previously
described
by
Corriu,
Andre,
Schott,
Aliehel
&
Stoclet
(1
994).
Briefly,
second
and
third
generation
miiesenter
ic
arteries
were
incubated
for
18
h
with
16
6
#tCi
myo-5-[3H]inositol
(ml
MEM)-'
at
37
°C
and
gassed
w,Nith
95%
02-5%
CO2.
When
required,
smnall
resistance
arteries
weie
incubated
with
600
ng
mnlF'
PTX
foi
12
h
at
37
'C.
Unincorporated
isotope
was
renmoved
by
w\ashing
with
PSS,
vessels
were
then
incubated
for
30
mnin
in
P88
cointaininig
10
mM
LiCl
kept
at
37
'C
and
gassed
continucously
with
a
iiixtuie
of
95%
02-5%
CO2
(pH
74).
After
washing,
vessels
were
stimulated
with
ATP
(1
and
3
mm)
fom
15
s,
rapidly
fiozen
uising
an
aluminium
clamp
pre-cooled
in
liquidl
nitrogen,
and
incubated
in
an
ice-cold
20%
trichloroacetic
acid
solution.
All
the
following
steps
were
performed
at
4
'C.
Vessels
wvere
ground
and
centrifuged
at
2000
g
for
10
min
at
4
'C,
the
supernatant
was
neutralized
to
pH
7
5
with
KOH
(10
N)
and
then
transferir
ed
to
columns
of
AG1
-
X8
anion
exchange
iesin.
Free
inositol
was
eluted
with
ice-cold
deionized
ater
(Millip)ore).
[3H]inositol
inonophosphate
anid
[3H]inositol
diphosphate
were
eluted
in
0
2
m
ammlloniulm
peCiiod
between
each
al)plication.
691
J.
Physiol.
492.3
foi-iiiate-0-
I
m
foi-iiiie
ael(i
an(I
0-4
Ai
aiii
iiioniuiii
foi-iiiate-0-
I
m

692
G.
J
L.
Lag
formic
acid,
respectively.
[3H]inositol
trisphosphate
was
eluted
in
1
M
ammoniuin
formate-Ol1
forinic
acid
and
counted.
Results
were
expressed
in
termns
of
disintegrations
per
minute
of
[3H]-
1P3
per
microgram
D.NA
(d.p.m.
(jg
DNA)-').
DNA
content
was
measured
as
described
by
Brunk,
Jones
&
Jones
(1979).
Expression
of
results
and
statistical
analysis
Contractions
were
expiessed
as
a
percentage
of
the
maximal
conitractile
response
obtained
with
10
#.-
noradrenaline
(NArnax).
The
contractile
response
to
10
AM
noradrenaline
was
not
significantly
different
between
control
and
PTX-treated
vessels
being
2-5
+
0-14
g
(ti
=
12)
and
2-47
+
0-17
g
(mean+
S.E.M.,
i
=
12),
respectively.
Therefore,
contractions
were
also
expressed
as
a
percentage
of
NAnmax
on
vessels
treated
with
PTX.
The
contraction
to
ATP
was
measured
at
the
peak
of
the
response.
The
change
in
[Ca2+]j
was
calculated
using
the
equation
previously
described
by
Grynkiewicz,
Poenie
&
Tsien
(1985).
The
sensitivity
of
the
vessels
to
purinergic
analogues
is
expressed
as
the
EC50
(the
concentration
of
agonist
required
to
give
the
half-maximal
response
of
the
vessels)
and
values
were
obtained
by
logit/log
regression
analy
sis.
A
A
[ATP]
(mM)
0
01
rau
0
0-1
id
and
others
J
Physiol.
492.3
[3H]IP3
content
is
expressed
as
d.p.m.
(,ug
DNA)-'.
All
results
are
expressed
as
means
+
S.E.M.
of
n
experiments.
Multivariate
analysis
of
variance
(MANOVA)
was
used
for
statistical
analysis
with
the
SYSTAT
program
(Evanson,
IL,
USA).
Means
were
considered
significantly
different
when
the
P
value
was
less
than
0
05.
Drugs
Acetvlcholine
chloride,
A.MP,
ADP/)S,
AT'SP
disodium
salt,
ADP,
a,,I-MeATP
lithium
salt,
caffeine,
noradrenaline
bitartrate,
NiCI2,
PTX,
Reactive
Blue
2,
thapsigargin
and
UTP
sodium
salt
were
purchased
from
Sigma
(Grenoble,
France).
myo-[3H]inositol
was
purchased
from
Amersham
Life
Science
(Amersham,
UK),
nitrendipine
and
suramin
were
a
generous
gift
from
Bayer
AG
(Wuppertal,
Germany
and
Paris,
France).
2-MeSATP
and
PPADS
sodium
salt
was
purchased
from
Research
Biochemicals
International
(Natick,
MA,
USA).
SK&F
96365
was
a
generous
gift
from
SmithKline
Beecham
Pharmaceuticals
(London,
UK).
Nitrendipine
was
dissolved
in
absolute
ethanol.
Thapsigargin
was
dissolved
in
dimethyl
sulphoxide
(1
mg
ml-')
and
the
final
concentration
of
dimethyl
sulphoxide
in
the
bath
was
0-1
%
All
other
drugs
were
diluted
in
deionized
water
(MIillipore).
1
Ca2+
LL00
nm
60
s
[L00
mg
60
s
13
free
PSS
+
0-5
mm
EGTA
T
3
mm
ATP
B
x
E
z
C
-
c
0
0
90
r
45
F
0
0-001
0.01
0.1
[ATP]
(mM)
1
3
Figure
1.
Effects
of
ATP
on
[Ca2+]i
and
contraction
A,
representative
traces
showing
effects
of
ATfl
on
the
change
in
[Ca2+]i
(upper
traces)
and
conttaction
(lower
traces)
of
the
small
mesenteric
artery
of
the
rat
in
nor-mal
and
in
Ca2+-free
medium.
B,
histograms
showvin1g
the
concentration-dependent
contractile
responses
to
ATP,
in
normal
P88
(O,
it
=
5)
anid
in
Ca2+-firee
inediuin
(U,
7
=
5).
Here
and
in
otlher
figuies
contractions
are
expressed
as a
percenttage
of
the
maximnal
contractile
response
induced
by
10
AIm
noradrenaline
(.NAmax);
values
are
means
+
SEM.
P
<
0
001,
significantly
different
fiom
responise
obtained
in
normal
P88.
T
-j-
r-z--,

Ca2+
and
ATP
in
resistance
arteries
RESULTS
Effect
of
ATP
in
normal
PSS
and
in
Ca2e-free
medium
Figure
1A
shows
typical
traces
for
the
change
in
[Ca2P]i
and
the
contraction
produced
by
ATP
(0
001-3
mM)
in
normal
and
in
Ca2+-free
PSS.
Figure
lB
shows
the
means
of
the
ATP-induced
contractions
in
normal
and
in Ca2+-free
medium,
expressed
as
a
percentage
of
the
contractile
response
obtained
with
noradrenaline
(10
/SM).
In
normal
PSS,
addition
of
ATP
induced
a
transient
increase
in
both
[Ca2+]i
and
tension
in
a
concentration-dependent
manner.
The
highest
ATP
concentration
used
in
this
study
was
3
mM.
Higher
concentrations
of
ATP
did
not
induce
greater
[Ca2P]i
or
contractile
responses.
In
Ca2+-free
PSS,
ATP
elicited
increases
in
both
[Ca2P]i
and
tension
only
at
a
concentration
of
3
mm.
The
maximal
response
of
the
vessel
to
ATP
(3
mM)
was
significantly
reduced
in
Ca2+-free
medium
(P
<
0
001).
Removal
of
the
endothelium
did
not
modify
either
the
maximal
response
or
the
sensitivity
of
the
vessels
to
ATP.
The
maximal
contraction
(%
NAmax)
and
EC50
(jUM)
values
of
ATP
were,
respectively,
76
+
7
%
and
280
+
105
/M
in
the
A
presence
of
endothelium
and
78
+
4%
and
225
+
44/SM
in
the
absence
of
the
endothelium.
ADP
produced
contraction
of
the
vessels
in
a
concentration-dependent
manner
with
the
same
potency
as
ATP
(as
shown
below
in
Fig.
8).
AMP
and
adenosine
were
not
able
to
produce
contraction
but
these
metabolites
relaxed
arteries
pre-contracted
by
noradrenaline
at
concentrations
higher
than
300
#M
(not
shown).
Maximal
responses
of
each of
these
agents
were
obtained
at
3
mm
and
were
40%
relaxation
NAmax
for
AMP
and
60%
relaxation
NAmax
for
adenosine.
Involvement
of
Ca2+
entry
and
release
from
intra-
cellular
stores
In
order
to
distinguish
the
Ca2+
entry
and
the
internal
Ca2+
release
components
of
the
contraction,
ATP
was
used
at
a
concentration
of
either
1
or
3
mM
in
the
following
experiments.
Figure
2A
shows
representative
traces
of
the
effect
of
ATP
(1
mM)
both
on
the
increase
in
[Ca2P],
and
on
the
contractile
responses
in
the
absence
or
presence
of
either
1
/SM
1
/SM
nitrendipine
S
~~~~~~~~~~~~~~~
~,.
-
30
uM
SK&F
96365
+
1
#M
nitrendipine
100
nlML
50
S
1__
I
__
°
__
200mgL
..
.
A
50
s..
ArM
4
-
.
A
TN
4
5.
s
1
MM
ATP
B
z
.-OI
0
0t
0
1
MM
ATP
1
MM
ATP
ITT
ttt
1
mM
ATP
Figure
2.
Effects
of
Ca2'-entry
blockers
on
[Ca2+]i
and
contraction
induced
by
ATP
A,
representative
traces
showing
the
influence
of
nitrendipine
(1
/sM)
and
SK&F
96365
(30
uM)
on
the
increase
in
both
[Ca2+],
(upper
traces)
and
tension
(lower
traces)
induced
by
1
mm
ATP.
B,
histograms
showing
the
contractions
induced
by
ATP
(1
mM)
in
the
absence
(O,
n
=
10)
and
in
the
presence
of
either
1
FM
nitrendipine
(U,
n
=
5),
30/uM
SK&F
96365
(0,
n
=
5)
or
SK&F
96365
plus
nitrendipine
(E,
n
=
10).
***
P
<
0
001,
significantly
different
from
the
control;
tt
P
<
0'01,
ttt
P
<
0o001,
significantly
different
from
the
contraction
produced
by
ATP
in
the
presence
of
nitrendipine;
ITT
P
<
0
001,
significantly
different
from
the
contraction
produced
by
ATP
in
the
presence
of
SK&F
96365.
...
tt
-s
jL,,ALAiL-
AL.j-.
J
Physiol.
492.3
693
4
-

J
Physiol.492.3
C.
J
L.
Lagaud
and
others
80
r
z
c
40
0
co
c)
0
1
mM
ATP
Figure
3.
Inhibition
by
nitrendipine
and
SK&F
96365
of
Ca2'-induced
contraction
in
ATP-
exposed
arteries
Histograms
showing
contractions
induced
by
addition
of
CaCl2
(3
mM)
in
small
mesenteric
resistance
arteries
of
the
rat
exposed
to
ATP
(1
mM)
in
Ca2P-free
medium
in
the
absence
(El,
n
=
12)
and
in
the
presence
of
either
1
/tM
nitrendipine
(U,
n
=
7)
or
30
AM
SK&F
96365
(n
=
5).
***
P<
0001,
significantly
different
from
control.
nitrendipine
or
1
,UM
nitrendipine
plus
30
,C4M
SK&F
96365.
ATP
(1
mM)
produced
a
transient
increase
in
both
[Ca2+]j
and
tension.
In
the
presence
of
nitrendipine
(1
,UM),
the
peak
responses
to
ATP
were
inhibited
by
50%.
Responses
to
ATP
(1
mM)
were
abolished
by
the
simultaneous
presence
of
nitrendipine
(1
,UM)
and
SK&F
96365
(30
,UM).
Figure
2B
shows
the
mean
contractile
responses
to
1
mm
A
ATP
with
nitrendipine
inhibiting
the
contractile
response
to
ATP
by
50%,
SK&F
96365
alone
by
82%
and
the
combination
of
nitrendipine
and
SK&F
96365
abolishing
the
contractile
response
to
ATP.
Experiments
were
also
performed
on
vessels
that
had
been
depleted
of
their
intracellular
Ca2P
stores
as
described
in
Methods
above.
Under
these
conditions,
small
mesenteric
100
nML
1
uM
thapsigargin
60s
200
m
gL
l
|
~~~~~~~3
mm
ATP
60
s
3
mM
ATP
10
mM
caffeine
3
mM
ATP
B
-
Iv
z
10
c
20
0
0~
C)
-
3
mM
ATP
Figure
4.
Effects
of
caffeine
and
thapsigargin
on
responses
to
ATP
in
Ca2e-free
medium
A,
representative
traces
showing
effects
of
caffeine
(10
mM)
and
thapsigargin
(1
#M)
on
the
increase
in
both
[Ca2+]i
(upper
traces)
and
tension
(lower
traces)
induced
by
ATP
(3
mM)
in
Ca2P-free
medium.
B,
histograms
showing
contractions
induced
by
ATP
(3
mM)
in
Ca2+-free
medium
in
the
absence
(LO,
n
=
10)
and
in
the
presence
of
either
10
mm
caffeine
(U,
n
=
5)
or
1
#tM
thapsigargin
(iJ,
n1
=
5).
*
P
<
005,
**
P
<
0
01,
significantly
different
from
control;
tt
P
<
0
01,
significantly
different
from
the
contraction
produced
by
ATP
in
the
presence
of
thapsigargin.
694
T
Ca2+
and
A
TP
in
resistance
arteries
arteries
maintained
in
CPa2-free
PSS
containing
2
mM
EGTA,
did
not
respond
to
ATP
alone
or
to
exogenous
application
of
3
mm
CaCl2
in
the
absence
of
ATP.
As
illustrated
in
Fig.
3,
addition
of
3
mm
CaCl2
in
the
presence
of
ATP
(1
mM)
produced
a
small
contractile
response
of
the
vessels
that
was
decreased
by
93%
by
nitrendipine
at
(1
,UM).
This
contractile
response
to
CaCl2
was
abolished
by
30
,UM
SK&F
96365.
In
Ca2+-free
medium,
caffeine
(10
mM)
produced
a
fast
transient
increase
in
[Ca2+]i
and
contraction
and
prevented
a
subsequent
response
to
ATP
(3
mM;
Fig.
4A).
Thapsigargin
(1
FM)
alone
did
not
induce
responses
but
reduced
the
increase
in
[Ca2+]i
and
tension
produced
by
3
mm
ATP
in
Ca2+-free
medium.
Figure
4B
shows
the
mean
of
the
contractile
response
to
ATP
(3
mM);
it
was
reduced
by
thapsigargin
(1
uM)
and
abolished
by
caffeine
(1O
mM)
in
Ca2+-free
medium.
The
caffeine
response
was
not
affected
by
prior
stimulation
with
either
ATP
or
thapsigargin
(data
not
shown).
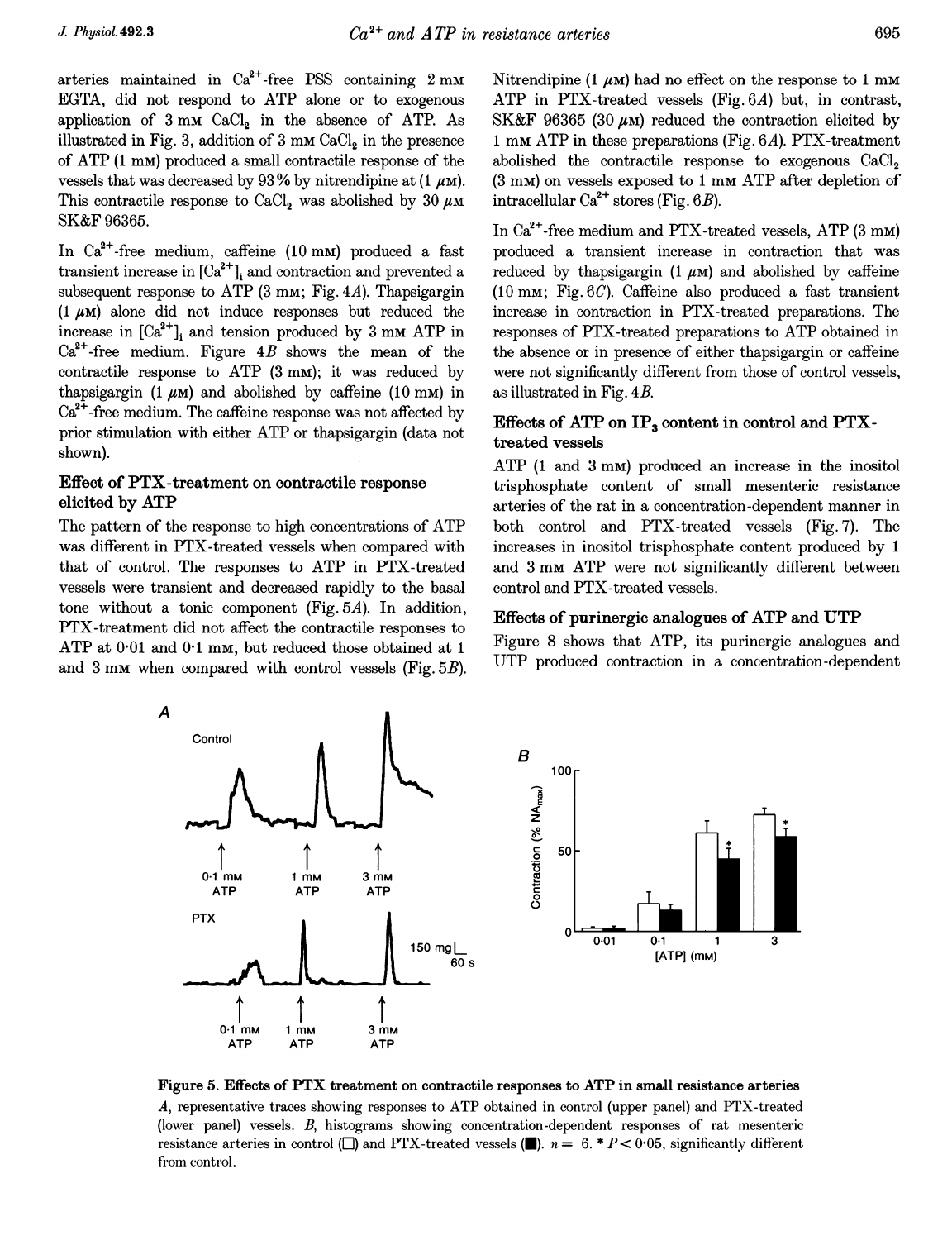
Effect
of
PTX-treatment
on
contractile
response
elicited
by
ATP
The
pattern
of
the
response
to
high
concentrations
of
ATP
was
different
in
PTX-treated
vessels
when
compared
with
that
of
control.
The
responses
to
ATP
in
PTX-treated
vessels
were
transient
and
decreased
rapidly
to
the
basal
tone
without
a
tonic
component
(Fig.
5A).
In
addition,
PTX-treatment
did
not
affect
the
contractile
responses
to
ATP
at
001
and
01
mm,
but
reduced
those
obtained
at
1
and
3
mm
when
compared
with
control
vessels
(Fig.
5B).
Nitrendipine
(1
FM)
had
no
effect
on
the
response
to
1
mM
ATP
in
PTX-treated
vessels
(Fig.
6A)
but,
in
contrast,
SK&F
96365
(30
FUM)
reduced
the
contraction
elicited
by
1
mM
ATP
in
these
preparations
(Fig.
6A).
PTX-treatment
abolished
the
contractile
response
to
exogenous
CaCl2
(3
mM)
on
vessels
exposed
to
1
mm
ATP
after
depletion
of
intracellular
Ca2+
stores
(Fig.
6B).
In
Ca2+-free
medium
and
PTX-treated
vessels,
ATP
(3
mM)
produced
a
transient
increase
in
contraction
that
was
reduced
by
thapsigargin
(1
FUM)
and
abolished
by
caffeine
(1O
mM;
Fig.
6C).
Caffeine
also
produced
a
fast
transient
increase
in
contraction
in
PTX-treated
preparations.
The
responses
of
PTX-treated
preparations
to
ATP
obtained
in
the
absence
or
in
presence
of
either
thapsigargin
or
caffeine
were
not
significantly
different
from
those
of
control
vessels,
as
illustrated
in
Fig.
4B.
Effects
of
ATP
on
IP3
content
in
control
and
PTX-
treated
vessels
ATP
(1
and
3
mM)
produced
an
increase
in
the
inositol
trisphosphate
content
of
small
mesenteric
resistance
arteries
of
the
rat
in
a
concentration-dependent
manner
in
both
control
and
PTX-treated
vessels
(Fig.
7).
The
increases
in
inositol
trisphosphate
content
produced
by
1
and
3
mm
ATP
were
not
significantly
different
between
control
and
PTX-treated
vessels.
Effects
of
purinergic
analogues
of
ATP
and
UTP
Figure
8
shows
that
ATP,
its
purinergic
analogues
and
UTP
produced
contraction
in
a
concentration-dependent
A
Control
01
mm
1m
01mM
1mM
3mM
ATP ATP
ATP
PTX
150
mgL
60
s
B
100
z
T
x~~~~~~~~~~~
C
50
0
C)
i5
c
0
0
0-01
0-1
1
3
[ATP]
(mM)
0-1
mM
1
mM
3mM
ATP
ATP
ATP
Figure
5.
Effects
of
PTX
treatment
on
contractile
responses
to
ATP
in
small
resistance
arteries
A,
representative
traces
showing
responses
to
ATP
obtained
in
control
(upper
panel)
and
PTX-treated
(lower
panel)
vessels.
B,
histograms
showing
concentration-dependent
responses
of
rat
inesenteric
resistance
arteries
in
control
([)
and
PTX-treated
vessels
(U).
n
=
6.
*
P
<
005,
significantly
different
from
control.
J
Physiol.
492.3
695

0.
J
L.
Lagaud
and
others
A
E
z
C:
0
0
cu
c)
1
mM
ATP
C
x
z
c
0
C
0
B
E
z
C:
0
(5
0
J
Physiol.492.3
100
r
50
0
1
mM
ATP
100
r
50
0
l'F--M
3
mM
ATP
Figure
6.
Effects
of
PTX-treatment
on
contractile
responses
to
ATP
in
small
resistance
arteries
A,
histograms
showing
the
contractions
induced
by
ATP
in
arteries
treated
with
PTX
in
the
absence
(l)
and
in
the
presence
of
either
1
uLM
nitrendipine
(NIT;
*)
or
30
,UM
SK&F
96365
(E).
n
=
5.
**
P
<
0
01,
significantly
different
from
the
control;
tt
P
<
001
significantly
different
from
the
contraction
produced
by
ATP
in
the
presence
of
nitrendipine.
B,
histograms
showing
contractions
induced
by
addition
of
CaCl2
(3
mM)
to
vessels
exposed
to
1
mm
ATP
in
Ca2+-free
medium
in
control
(El,
n
=
12)
and
600
ng
ml-'
PTX-treated
preparations
(U,
n
=
5).
***
P
<
0001,
significantly
different
from
control.
C,
histograms
showing
the
contractions
induced
by
3
mm
ATP
in
arteries
treated
with
PTX
in
Ca2+-free
medium
in
the
absence
(EO,
n
=
10)
and
in
the
presence
of
either
10
mm
caffeine
(U,
n
=
5)
or
1
,UM
thapsigargin
(lM,
n
=
5).
*
P
<
0
05,
**
P
<
0
01,
significantly
different
from
control;
tt
P
<
001,
significantly
different
from
the
contraction
produced
by
ATP
in
the
presence
of
thapsigargin.
manner
with
an
order
of
potency
of
ac,,-MeATP>
2-
MeSATP
>
ATP
=
ADP
>
UTP.
The
maximal
contraction
(%)
and
EC50
(#UM)
values
of
each
of
these
agents
were,
respectively:
75
+
5%
and
0
7
+
0
4
/M
for
z,/-MeATP;
65
+
7
%
and
60
+
25
uM
for
2-MeSATP;
76
+
7
%
and
280+105
#M
for
ATP;
75+8%
and
254+50/SM
for
ADP;
113
+
17%
and
79
6
/M
for
UTP
(n=
5
for
all
experiments
except
UDP
where
n
=
6).
Removal
of
A
z
cn
CD
ci
V6
a.
0
1
[ATP]
(mM)
endothelium
did
not
modify
the
concentration-response
curves
for
ATP
and
its
analogues
(not
shown).
Also,
no
cross
desensitization
has
been
observed
between
ATP
and
UTP
in
mediating
contraction.
ATP
(3
mM)
was
able
to
produce
a
17
+
5-8%
increase
in
contraction
after
maximally
active
stimulation
of
the
vessels
with
UTP
(3
mM)
which
by
itself
produced
sustained
contraction
with
maximum
being
76
+
3
4%
of
the
maximal
contraction
B
80
*
40
*
....
...
_
..............:
0
1
3
[ATP]
(mM)
Figure
7.
Increases
in
IP3
content
induced
by
ATP
in
the
absence
(A)
and
presence
(B)
of
PTX
Histograms
showving
the
IP3
content
of
the
small
mesenteric
artery
of
the
rat
in
the
absence
(El,
n
=
7)
or
in
the
presence
of
1
mm
ATP
(ii1,
n
=
3)
or
3
IM
ATP
(U,
n
=
4)
in
control
(A)
and
PTX-treated
(B)
vessels,
respectively.
*
P<
0
05,
***
P<
0
001,
significantly
different
from
the
basal
level.
696
T

Ca2+
and
ATP
in
resistance
arteries
100
E
z
0
00
0
c
0
0.01
1
100
[Agonist]
(aM)
Figure
8.
Effects
of
purinergic
agonists
Concentration-response
curves
to
a,f6-MeATP
(0),
2-MeSATP
(A),
ATP
(O),
ADP
(O)
and
UTP
(0)
in
small
mesenteric
resistance
arteries
of
the
rat.
n
=
6.
(n
=
8).
Conversely,
UTP
(3
mM)
was
able
to
produce
a
75
+
2
1
%
increase
in
contraction
after
stimulation
of
the
vessels
with
ATP
(3
mM)
which
by
itself
produced
a
transient
increase
in
tension
with
a
peak
value
of
77
+
4
2%
of
the
maximal
contraction
(n
=
8).
Thus,
no
cross-desensitization
was
observed
for
ATP
and
UTP
in
producing
contraction.
A
x
E
z
-
0
0
._o
0
0
C
0
4
80
5
:
z
.-O
c
40
0
0
cJ
0
0
0
10
A"
***
***
T,I
0
n
0-01
0.1
[ATP]
(mM)
Effects
of
P2-purinoceptor
antagonists
As
illustrated
in
Fig.
9A,
suramin
(10-100
,UM)
abolished
the
contractile
responses
to
low
concentrations
of
ATP
(i.e.
less
than
0
1
mM)
and
reduced,
in
a
concentration-
dependent
manner,
the
contraction
elicited
by
high
concentrations
of
ATP.
Reactive
Blue
2
(30-100
,uM)
had
no
effect
on
contractions
induced
by
ATP
(Fig.
9B).
PPADS
B
D
0-1
[ATP]
(mM)
80
40
0
0-01
0-1
[ATP]
(mM)
0-1
[ATP]
(mM)
Figure
9.
Effects
of
P2-purinoceptor
antagonists
on
contractile
responses
to
ATP
Concentration-response
curves
of
rat
mesenteric
resistance
arteries
to
applications
of
ATP,
in
the
absence
(0)
and
in
the
presence
of
the
following
drugs.
A,
suramin
at
10
(0),
30
(A),
60
(A)
and
80
/M
(E).
B,
Reactive
Blue
2
at
30
(0),
50
(A)
and
100
/AM
(A).
C,
PPADS
at
10
(0),
30
(A)
and
60
/LM
(A).
D,
a,/3-
MeATP
at
10
/M
(0).
n
=
5.
*
P<
0
05,
**
P<
0
01,
***
P<
0
001,
significantly
different
from
control.
J
Physiol.
492.3
697
on
_
80r
1
1
1

G.
J
L.
Lagaud
and
others
J
Physiol.492.3
B
100
r
T
50
10
10
[Agonist]
(uM)
0
)0
1000
300
#M
ADP/S
Figure
10.
Pharmacological
characterization
of
P2Y-purinoceptors
in
the
small
mesenteric
artery
of
the
rat
A,
concentration-response
curves
for
2-MeSATP
(A),
ATP
(>)
and
ADP
(O)
obtained
in
the
presence
of
60
,UM
PPADS.
B,
histograms
showing
responses
to
300
/Um
ADP/S
in
normal
PSS
(LI),
in
Ca2P-free
medium
(U),
in
normal
PSS
in
the
presence
of
60,UM
PPADS
(1)
or
100
/UM
Reactive
Blue
2
(1).
*
P
<
005,
**
P
<
001,
significantly
different
from
control;
tt
P
<
001,
significantly
diffierent
from
response
obtained
in
the
presence
of
PPADS.
n
=
5
in
all
experiments.
(10-60
/LM)
abolished
the
response
to
low
concentrations
of
ATP
and
inhibited
contractions
elicited
by
high
concentrations
of
ATP
(1
and
3
mM)
in
a
concentration-
dependent
manner
(Fig.
9C).
A
higher
concentration
of
PPADS
(100
/M)
did
not
produce
greater
inhibition
of
ATP
A
E
z
c
0
C.)
C
0
C
E
z
-0
C
0
t.g
0
cu
C
0
90
45
0
90
45
0
responses
(data
not
shown).
a,,f-MeATP
(10
JM)
abolished
responses
to
low
concentrations
of
ATP
(Fig.
9D).
A
very
small
component
resistant
to
the
desensitization
of
a,/3-
MeATP
(10
AM)
remained
at
1
and
3
mm
ATP.
B
100
1000
10
100
[UTP]
(uM)
[UTP]
(#lM)
D
100
1000
10
100
[UTP]
(#M)
[UTP]
(um)
1000
1000
Figure
11.
Effects
of
P2-purinoceptor
antagonists
on
contractile
response
to
UTP
Concentration-response
curves
of
rat
mesenteric
resistance
arteries
to
UTP,
in
the
absence
(0)
and
in
the
presence
of
the
following
drugs.
A,
suramin
at
10
(0),
30
(A),
60
(-)
and
80
/AM
(O).
B,
Reactive
Blue
2
at
30
(0),
50
(A)
and
100
um
(A).
C,
PPADS
at
10
(0),
30
(A)
and
60
#M
(A).
D,
x,/-MeATP
at
10
#tm
(0).
n
=
5.
698
A
100
x
E
z
c
50
.0
0
0

Ca2+
and
A
TP
in
resistance
arteries
Figure
1
OA
shows
concentration-contractile
response
curves
to
2-MeSATP,
ATP
and
ADP
obtained
after
blockade
of
the
P2X-purinoceptors
by
maximally
active
concentration
of
PPADS
(60
,uM).
ATP
and
its
purinergic
analogues
produced
contraction
in
a
concentration-dependent
manner
with
an
order
of
potency
of
2-MeSATP>
ATP
=
ADP.
The
maximal
contraction
(%)
and
EC50
(/SM)
values
of
each
of
these
agents
were,
respectively:
83
+
7%
and
33
+
7
,CM
for
2-MeSATP;
34
+
4%
and
933
+
100
,UM
for
ATP;
23
+
6%
and
1403
+
324
/tM
for
ADP
(n
=
5).
Contractions
produced
by
ATP
and
purinergic
analogues
obtained
in
the
presence
of
PPADS
(60
/uM)
were
abolished
by
Reactive
Blue
2
(100
,uM)
(not
shown).
The
contractile
response
produced
by
the
selective
P2Y-purinoceptor
agonist,
ADP/IS,
used
at
a
concentration
of
300
/M
was
not
significantly
affected
in
the
presence
of
PPADS
(60
,UM)
but
it
was
almost
completely
abolished
in
the
presence
of
Reactive
Blue
2
(100
,UM)
(Fig.
1
OB).
In
addition,
50%
of
the
response
to
ADP/JS
could
be
obtained
in
Ca2+-free
medium
(Fig.
lOB).
As
illustrated
in
Fig.
11,
contractile
responses
to
UTP
were
not
affected
by
suramin,
Reactive
Blue
2,
PPADS
or
,,-
MeATP.
DISCUSSION
The
aim
of
the
present
study
was
to
investigate
the
relationship
between
the
stimulation
of
ATP
receptors,
the
increase
in
intracellular
calcium
and
contraction
in
the
small
mesenteric
artery
of
the
rat.
The
results
show
that
the
mechanisms
of
ATP-induced
increases
in
[Ca2+]j
and
contraction
vary
with
the
agonist
concentrations
and
receptor
subtypes.
At
concentrations
lower
than
1
mm,
ATP,
via
the
activation
of
P2X-purinoceptors,
produced
an
influx
of
calcium
through
nitrendipine-sensitive
and
SK&F
96365-sensitive
mechanisms.
These
mechanisms
are
believed
to
act
through
voltage-dependent
and
receptor-
operated
calcium
channels,
respectively.
At
concentrations
of
1
and
3
mm,
ATP
activated
P2Y-purinoceptors
and
induced
both
an
influx
of
calcium
sensitive
to
nitrendipine
via
a
PTX-sensitive
G
protein
and
the
release
of
Ca2+
from
an
internal
source
via
the
production
of
IP3.
In
the
small
mesenteric
artery
of
the
rat,
the
presence
of
the
endothelium
did
not
affect
the
contraction
produced
by
ATP.
In
addition,
no
endothelium-dependent
relaxant
effect
of
ATP
has
been
reported
in
the
same
type
of
artery
(Juul,
Plesner
&
Aalkjaer,
1993).
By
investigating
the
effect
of
ATP
metabolites,
ATP
and
ADP
were
found
to
be
equipotent
in
eliciting
contraction.
AMP
and
adenosine
were
not
able
to
induce
contraction
but
rather
they
relaxed
pre-
contracted
arteries
at
high
concentrations
(>
300
/,M).
Thus,
it
is
unlikely
that
the
mechanisms
by
which
ATP
induces
the
increase
in
[Ca2+]i
and
contraction
involved
the
effect
of
its
metabolites
such
as
AMP
or
adenosine
even
though
the
presence
of
ectonucleotidases
on
the
membrane
of
rat
Plesner,
1991)
probably
leads
to
an
underestimation
of
ATP
potency.
In
the
present
study,
low
concentrations
of
ATP
(<
1
mM)
induced
calcium
entry
whereas
higher
concentrations
of
ATP
(3
mM)
produced
an
additional
release
of
calcium
from
internal
sources.
The
mechanisms
of
ATP-induced
calcium
entry
have
been
investigated
using
nitrendipine,
an
inhibitor
of
voltage-dependent
calcium
channels
and
SK&F
96365,
a
drug
described
as
a
selective
inhibitor
of
receptor-
operated
calcium
channels
in
many
cell
types
such
as
human
platelets,
neutrophils,
PC12
cells
and
endothelial
cells
(Fasolato,
Pizzo
&
Pozzan,
1990;
Mlerritt
et
at.
1990).
The
increase
in
both
[Ca2+]i
and
contraction
at
low
concentrations
of
ATP
(<
1
mM)
were
abolished
by
nitrendipine.
Responses
obtained
with
1
mm
ATP
were
inhibited
by
50%
with
nitrendipine
and
the
nitrendipine-
insensitive
component
was
abolished
by
SK&F
96365.
In
addition,
the
contractile
response
to
exogenous
Ca2+
of
ATP-exposed
vessels
was
inhibited
by
both
nitrendipine
and
SK&F
96365.
SK&F
96365
has
been
shown
to
inhibit
both
KCl-induced
contractions
and
the
nifedipine-
insensitive
component
of
contractions
induced
by
acetylcholine,
9,11
-dideoxy-9z,11x-methanoepoxy
prosta-
glandin
F2a
(U46619)
or
leukotriene
D4
on
guinea-pig
trachea
(Cuthbert,
Gardiner,
Nash
&
Poll,
1994).
Indeed,
SK&F
96365
was
able
to
abolish
the
contractile
response
induced
by
KCl
depolarization
which
produced
calcium
entry
via
voltage-operated
calcium
channels
in
the
small
mesenteric
artery
of
the
rat
(data
not
shown).
It
has
a
dual
effect
on
the
calcium
entry
through
inhibition
of
the
opening
of
both
receptor-operated
and
voltage-dependent
calcium
channels.
It
has
been
reported
that
SK&F
96365
does
not
block
the
contractile
effect
of
ATP
in
rabbit
ear
artery
(Merritt
et
at.
1990).
However,
purinoceptor
subtypes
activated
by
ATP
in
rabbit
ear
artery
showed
similarities
and
differences
from
data
obtained
in
other
smooth
muscle
preparations
regarding
the
relative
potencies
of
P2x-purinoceptor
agonists
(for
review
see
Kennedy
&
Leff,
1995).
Thus,
it
is
possible
that
the
ATP-
gated
channels
of
rabbit
ear
artery
differ
from
those
activated
in
rat
small
mesenteric
arteries
explaining
the
differential
sensitivity
of
the
channel
to
SK&F
96365.
Nevertheless,
the
results
obtained
here
with
nitrendipine
and
SK&F
96365
support
the
view
that
ATP
promotes
an
influx
of
Ca2P
through
both
voltage-dependent
calcium
channels
and
receptor-operated
channels
sensitive
to
SK&F
96365
in
the
small
mesenteric
resistance
artery
of
the
rat.
These
data
are
in
accordance
with
those
reported
in
the
literature
showing
that
ATP
elicits
contraction
by
activating
a
non-selective
cationic
channel,
permeable
to
Na+
and
Ca2+,
that
can
have
dual
excitatory
functions,
depolarizing
the
cell
leading
to
the
opening
of
voltage-
dependent
Ca2+
channel,
and
functioning
as
a
direct
calcium
entry
pathway
(Suzuki,
1985;
Benhain
&
Tsien,
mesenteric
resistance
arteries
(Juul,
Liischer,
Aalkjeer
&
J:
Physiol.
492.3
699
1987;
Brake
et
al.
1994;
Valera
et
al.
1994).

G.
J
L.
Lagaud
and
others
The
present
report
also
shows
that
in
addition
to
inducing
calcium
entry,
maximally
active
concentrations
of
ATP
(i.e.
3
mM)
produced
an
increase
in
[Ca2+]i
and
an
arterial
contraction
in
Ca2+-free
medium.
Moreover,
high
concentrations
of
ATP
(1
and
3
mM)
induced
an
increase
in
IP3
content
of
small
mesenteric
arteries.
Taken
together,
these
results
suggest
that,
in
these
arteries,
ATP
activates
the
formation
of
IP3,
probably
through
the
phospholipase
C
pathway,
which,
in
turn,
elicits
the
release
of
Ca2+
from
intracellular
stores
located
in
the
sarcoplasmic
reticulum.
Although
ATP
produced
a
slight
increase
of
IP3
content
at
a
concentration
of
1
mm,
it
was
not
able
to
elicit
an
increase
in
either
[Ca2+],
or
a
contraction
in
Ca2+-free
medium
and
in
normal
PSS
after
blockade
of
calcium
entry
with
nitrendipine
and
SK&F
96365.
The
latter
observations
support
the
view
that
the
increase
in
IP3
content
elicited
by
1
mM
ATP
might
not
be
sufficient
to
elicit
the
release
of
Ca2+
from
internal
source,
whereas
it
was
sufficient
when
the
arteries
were
exposed
to
3
mm
ATP.
The
above
data
are
in
accordance
with
those
reported
by
different
authors
within
either
isolated
aorta
or
aortic
smooth
muscle
cells
of
the
rat
(Nambi,
Hsiao-Ling
&
Nambi,
1990;
Kitajima,
Ozaki
&
Karaki,
1993).
However,
the
present
results
contrast
with
those
in
rat
portal
vein
showing
that
ATP
released
Ca2+
from
intracellular
stores
by
a
mechanism
that
involved
Ca2+
entry
(Pacaud,
Gregoire
&
Loirand,
1994).
It
is
unlikely
that
the
latter
mechanism
occured
within
small
mesenteric
arteries
since
ATP
was
still
able
to
induce
an
increase
in
both
[Ca2+]i
and
a
contraction
in
Ca2+-free
medium.
It
should
be
noted
that
the
absence
of
extracellular
calcium
did
not
result
in
lowering
the
concentration
of
ATP
required
to
produce
the
release
of
calcium
from
intra-
cellular
stores.
In
addition,
responses
to
ATP
(i.e.
3
mM)
were
not
significantly
different
in
Ca2+-free
medium
or
in
normal
PSS
in
the
presence
of
nitrendipine.
Obviously,
it
is
not
possible
to
determine
whether
or
not
the
effect
of
low
concentrations
of
ATP
(<
1
mM)
was
modified
as
a
consequence
of
altered
affinity
since
these
responses
were
entirely
mediated
by
calcium
entry.
However,
the
response
upon
addition
of
Ca2+
to
vessels
exposed
to
ATP
(1
mM)
in
Ca2'-free
medium
was
identical
to
that
elicited
by
the
same
concentration
of
ATP
in
Ca2+-containing
medium.
The
nature
of
the
intracellular
stores
released
by
ATP
through
the
1P3
pathway
was
studied
in
the
small
mesenteric
arteries
of
the
rat.
In
Ca2+-free
medium,
caffeine,
an
activator
of
Ca2+-induced
Ca2+
release
mechanism,
induced
a
fast
transient
increase
in
[Ca2+]i
and
contraction
and
completely
abolished
responses
to
ATP
(3
mM).
By
contrast,
responses
to
caffeine
were
not
affected
by
prior
stimulation
with
3
mm
ATP.
Also
in
Ca2+-free
medium,
thapsigargin
(1
uM),
an
inhibitor
of
the
sarcoplasmic
reticulum
Ca2+-
ATPase,
had
no
effect
on
either
[Ca2+]i
or
contraction
when
added
to
unstimulated
vessels,
but
it
decreased
by
65%
the
Andriantsitohaina
et
al.
1995).
The
results
suggest
that
two
intracellular
Ca2+
pools
releasable
by
ATP
occur
in
these
resistance
arteries
which
are
both
caffeine
sensitive,
but
only
one
of
them
is
sensitive
to
thapsigargin.
It
cannot
be
ruled
out
that
other
Ca2+-ATPases
which
are
insensitive
to
thapsigargin
might
exist
in
the
small
mesenteric
resistance
artery.
The
latter
might
explain
the
effectiveness
of
thapsigargin
in
inducing
contraction
by
itself
in
Ca2+-free
medium
and
in
depleting
the
intracellular
Ca2P
pools
sensitive
to
ATP.
Indeed,
thapsigargin
has
been
found
to
inhibit
the
100
kDa
but
not
other
forms
of
sarcoplasmic
reticulum
Ca2+-ATPases
(Papp
et
al.
1991).
Finally,
caffeine
releases
another
independent
intracellular
Ca2+
pool
which
is
insensitive
to
ATP
and
thapsigargin.
These
results
do
not
fit
with
the
model
proposed
by
Tatsuro,
Lino,
Yamazawa
&
Endo
(1991),
obtained
from
rat
portal
vein,
showing
that
all
intracellular
Ca2+
pools
are
sensitive
to
IP3
and
only
one
of
them
is
sensitive
to
caffeine.
The
differential
results
can
be
attributed
to
structural
and
functional
heterogeneity
of
sarcoplasmic
reticulum
between
vascular
preparations
(Low,
Kwan
&
Daniel,
1992;
Tribe,
Borin
&
Blaustein,
1994).
The
possible
implication
of
a
G
protein
in
the
mechanism
of
action
of
ATP
in
rat
resistance
arteries
was
investigated
by
studying
the
effect
of
PTX
which
has
been
shown
to
ADP-
ribosylate
a
40-41
kDa
membrane
protein
identified
as
a
subunit
of
a
Gi
protein
in
these
resistance
arteries
(Andriantsitohaina
et
al.
1990).
PTX
did
not
inhibit
contractions
produced
by
noradrenaline
suggesting
that
firstly
it
did
not
alter
the
contractile
capacity
of
the
vessel
and
secondly
contractile
responses
to
noradrenaline
did
not
involve
the
activation
of
PTX-sensitive
G
protein.
However,
PTX
inhibited
contractions
produced
by
high
concentrations
of
ATP
(i.e.
1
and
3
mM)
but
not
those
of
low
concentrations.
Also,
PTX
treatment
did
not
inhibit
contractile
responses
to
ATP
(3
mM)
in
Ca2+-free
medium.
Moreover,
the
increases
in
IP3
content
of
the
vessels
produced
by
ATP
were
not
affected
by
PTX
treatment.
These
results
suggest
that
the
response
to
ATP
acting
through
the
activation
of
phospholipase
C/IP3
pathway
was
not
linked
to
a
PTX-sensitive
G
protein.
By
contrast,
responses
to
exogenous
calcium
in
vessels
which
were
exposed
to
ATP,
after
having
previously
depleted
their
intracellular
calcium
stores,
were
abolished
by
PTX.
Furthermore,
PTX
abolished
the
inhibitory
effect
of
the
voltage-dependent
calcium
entry
blocker,
nitrendipine,
but
not
that
of
the
receptor-operated
calcium
entry
blocker,
SK&F
96365,
on
responses
to
ATP.
Taken
together,
these
results
suggest
that
a
PTX-sensitive
G
protein
is
involved
in
the
external
Ca2+-dependent
contraction
produced
by
ATP
in
small
mesenteric
arteries
of
the
rat.
The
calcium
entry
component
of
ATP-induced
responses
affected
by
PTX
involved
the
activation
of
nitrendipine-sensitive
influx.
Interestingly,
such
a
mechanism
has
been
reported
regarding
the
effect
of
neuropeptide
Y
within
the
same
type
of
artery
(Andriantsitohaina
et
at.
1990).
responses
elicited
by
ATP
(3
mM)
as
previously
reported
by
J
PhysioL
492.3
700

Ca2+
and
A
TP
in
resistance
arteries
The
above
data
show
that
ATP
mediates
the
increase
in
[Ca2+]i
and
contraction
through
several
transduction
pathways
in
rat
small
mesenteric
arteries.
Therefore,
the
hypothesis
that
ATP
mediates
its
effect
through
activation
of
multiple
receptor
subtypes
coupled
to
different
signal
transduction
mechanisms
was
tested
using
several
purinoceptor
agonists
and
antagonists.
Pharmacological
characterization
of
purinoceptors
shows
that
P2x-purinoceptors,
which
are
members
of
ligand-gated
channels
(Brake
et
al.
1994;
Valera
et
al.
1994),
are
involved
in
the
contractile
response
induced
by
low
concentrations
of
ATP
in
the
small
mesenteric
artery
of
the
rat.
This
conclusion
is
based
on
the
following
observations.
(1)
ATP,
at
concentrations
lower
than
1
mM,
stimulated
a
receptor-operated
channel
sensitive
to
SK&F
96365.
ATP
and
its
purinergic
analogues
induced
concentration-dependent
contraction
of
rat
small
mesenteric
arteries.
(2)
The
rank
order
of
potency
of
these
agonists
(a.,fl-MeATP
>
2-MeSATP
>
ATP
=
ADP)
classifies
this
receptor
as
a
P2x-purinoceptor
subtype
(Burnstock
&
Kennedy,
1985).
(3)
Suramin,
a
P2-purinoceptor
antagonist
without
any
preference
for
P2X-
and
P2Y-subtypes
(Burnstock
&
Warland,
1987),
inhibited
the
response
to
ATP
in
a
concentration-dependent
manner.
At
a
concentration
of
80
/SM,
suramin
abolished
the
response
to
ATP.
(4)
Reactive
Blue
2
used
in
the
range
of
concentrations
(30-100
/tM)
at
which
it
preferentially
inhibits
P2Y-purinoceptors
(Burnstock
&
Warland,
1987),
did
not
affect
the
contractile
response
to
ATP.
(5)
The
newly
described
selective
P2X-purinoceptor
antagonist,
PPADS,
inhibited
the
contractile
response
to
ATP
in
a
concentration-dependent
manner.
Although
it
is
difficult
to
decide
whether
or
not
the
antagonism
is
competitive,
since
maximum
effect
of
ATP
had
not
been
reached
in
the
presence
of
PPADS.
The
extent
of
the
inhibitory
effect
of
PPADS
on
ATP
response
was
identical
to
that
obtained
in
rabbit
ear
artery
and
saphenous
vein
after
activation
of
P2x-purinoceptors
by
a,,f-MeATP
(Ziganshin,
Hoyle,
Lambrecht,
Mutschler,
Biiumert
&
Burnstock,
1994).
Finally,
desensitization
of
P2X-purinoceptors
by
a,,-
MeATP
abolished
the
response
to
ATP.
It
has
to
be
noted
that
none
of
the
antagonists
used
in
the
present
study
affected
contractile
responses
to
UTP.
Furthermore,
no
cross-desensitization
has
been
observed
between
ATP
and
UTP
in
mediating
contraction
in
accordance
with
data
reported
by
Juul,
Plesner
&
Aalkjaer
(1992)
in
the
same
type
of
artery.
These
results
suggest
that
ATP
does
not
mediate
its
action
through
the
activation
of
P2U-
purinoceptors
even
though
rat
small
mesenteric
resistance
arteries
do
contain
P2U-purinoceptors.
In
addition,
the
above
results
also
show
that
ATP
at
high
concentrations
(i.e.
1
and
3
mM),
can
mediate
its
action
through
a
mechanism
that
corresponds
to
another
class
of
receptors
(i.e.
the
activation
of
a
phospholipase
C,/IP3
data have
shown
that
activation
of
sites
resembling
the
P2X-purinoceptor
could
be
linked
to
a
metabotropic
mechanism.
Moreover,
the
specific
P2X-purinoceptor
agonist,
a,,-MeATP
did
not
produce
an
increase
in
[Ca2+]i
or
contraction
of
small mesenteric
resistance
in
Ca2+-free
medium.
Therefore,
it
is
most
likely
that
P2-purinoceptors
other
than
P2X-purinoceptors
are
also
involved
in
the
mechanism
of
action
of
ATP
in
these
resistance
vessels.
Indeed,
after
blockade
of
P2X-purinoceptors
with
a
maximally
active
concentration
of
PPADS,
ATP
and
its
analogues
induced
concentration-dependent
contractions
of
rat
mesenteric
resistance
arteries
with
a
rank
order
of
potency
(2-MeSATP
>
ATP
=
ADP)
consistent
with
the
activation
of
P2Y-purinoceptor
subtypes
as
classified
by
Burnstock
&
Kennedy
(1985)
and
Fredholm
et
al.
(1994).
Under
these
experimental
conditions,
contractions
produced
by
ATP
and
its
analogues
were
abolished
by
Reactive
Blue
2.
In
addition,
the
contractile
responses
obtained
with
the
specific
P2Y-purinoceptor
agonist,
ADP/.?S,
were
not
affected
by
PPADS
but
were
abolished
by
Reactive
Blue
2.
Moreover,
ADP,8S
was
able
to
produce
contraction
in
Ca2+-free
medium
by
contrast
to
the
specific
P2x-purinoceptor
agonist
azf-MeATP
in
these
resistance
vessels.
Taken
together,
the
involvement
of
G
proteins
in
the
mechanisms
of
action
of
ATP,
the
rank
order
of
agonist
potency
after
blockade
of
P2X-purinoceptors
with
PPADS
and
the
effect
of
ADP/IS,
one
can
conclude,
that
ATP
stimulates
P2Y-purinoceptors
in
the
rat
small
mesenteric
resistance
artery.
The
exact
identity
of
the
P2Y-
purinoceptors
involved
remains
to
be
established.
In
conclusion,
the
present
findings
show
that
ATP
elicits
contraction
of
the
small
mesenteric
artery
of
the
rat
by
stimulating
at
least
two
types
of
P2-purinoceptor.
Low
concentrations
of
ATP
activate
P2x-purinoceptors
linked
to
a
ligand-gated
channels
that
can
have
dual
excitatory
functions,
depolarizing
the
cell
leading
to
the
opening
of
voltage-dependent
Ca2+
channel,
and
functioning
as
a
direct
calcium
entry
pathway.
High
concentrations
of
ATP
activate
P2Y-purinoceptors
one
of
which
is
coupled
to
PTX-
sensitive
G
protein
triggering
calcium
entry
via
dihyropyridine-sensitive
mechanism,
the
other
being
coupled
to
PTX-insensitive
G
protein
associated
with
phospholipase
C/IP3
pathway.
ANDRIANTSITOHAINA,
R.,
ANDRE,
P.
&
STOCLET,
J.
C.
(1990).
Pertussis
toxin
abolishes
the
effect
of
neuiopeptide
Y
on
rat
resistance
arteriole
contraction.
American
Journal
of
Physiology
259,
H1427-1432.
ANDRIANTSITOHAINA,
R.,
LAGAUD,
G.
J.-L.,
ANDRE,
A.,
MULLER,
B.
&
STOCLET,
J.
C.
(1995).
Effects
of
cGMP
on
calcium
handling
in
ATP-stimulated
rat
resistance
arteries.
American
Journal
of
Physiology
268,
H1223-1231.
BARNARD,
A.
E.,
BURNSTOCK,
G.
&
WEBB,
E.
T.
(1994).
G
protein-
coupled
receptors
for
ATP
and
other
nucleotides:
a
new
receptor
family.
Trends
in
Pharmacological
Sciences
15,
67-70.
pathway
and
a
PTX-sensitive
G
protein).
Up
to
now,
no
J
Physiol.
492.3
701

702
BENHAM,
C.
D.
(1989).
ATP-activated
channels
gate
calcium
entry
in
single
smooth
muscle
cells
dissociated
from
rabbit
ear
artery.
Journzal
of
Physiology
419,
689-701.
BENHANI,
C.
D.
&
TsIEN,
R.
AV.
(1987).
A
novel
receptor-operated
Ca
channel
activated
by-
ATP
in
smooth
muscle.
Xature
328,
275-278.
BRAKE,
A.
J.,
WAGENBACH,
M.
J.
&
JULIUS,
D.
(1994).
New
structulal
motif
for
ligand-gated
ion
channels
defined
by an
ionotropic
ATP
receptor.
Nature
371,
519-523.
BRUNK,
C.
F.,
JONES,
K.
C.
&
JONES,
T.
W.
(1979).
Assay
of
nanogram
quantities
of
DNA
in
cellular
homogenates.
A
nalytical
Biochemistry
92,
497-500.
BUKOSKI,
R.
D.,
BERGMANN,
C.,
GAIRARD,
A.
&
STOCLET,
J.
C.
(1989).
Intracellular
Ca1+
and
force
deteimined
simultaneously
in
isolated
resistance
arteries.
A
nerican
Journial
of
Physiology
257,
H1
728-1735.
BURNSTOCK,
G.
&
KENNEDY,
C.
(1985).
Is
there
a
basis
for
distinguishing
tw-o
types
of
P2-purinoceptor?
Ge
neral
Pharnmacology
16,
433-440.
BURNSTOCK,
G.
&
\V\ARLAND,
J.
J.
I.
(1987).
P1-purinoceptors
two
subtypes
in
the
rabbit
mnesenteric
artery:
reactive
blue
2
selectively
inhibits
responses
mediated
via
P2Y-
but
not
P2X-purinoceptor.
British
Journal
of
Pharmtiacology
90,
383-391.
CORRIU,
C.,
ANDRE,
P.,
SCHOTT,
C.,
MIlCHEL,
AI.
&
STOCLET,
J.
C.
(1994).
ANG
II
receptor
expression
and
function
during
phenotypic
modulation
of
rat
aortic
smooth
muscle
cells.
A
merican
Journial
of
Physiology
266,
H631-636.
CRACK,
B.
E.,
BEUKERS,
Al.
NV.,
AMcKECHNIE,
K.
C.
WV.,
IJZERMIANN,
A.
P.
&
LEFF,
P.
(1994).
Plharmacological
analysis
of
ecto-ATPase:
evidence
for
combined
enzy-me
inhibition
and
receptor
antagonism
in
P2X-
purinoceptor
ligands.
British
Journtal
of
Pharmnacology
113,
1432-1438.
CUTHBERT,
N.
J.,
GARDINER,
P.
J.,
NASH,
K.
&
POLL,
C.
T.
(1994).
Roles
of
Ca1+
influx
and
intracellular
Ca1+
release
in
agonist-
induced
contractions
in
guinea
pig
trachea.
American
Jourtnal
of
Physiology
266,
L620-627.
DUBYAK,
G.
R.,
COWEN,
D.
S.
&
AIEULLER,
L.
Al.
(1988).
Activation
of
inositol
phospholipid
breakdown
in
HL-60
cells
by
P2-purinergic
receptors
for
extracellular
ATP.
Journal
of
Biological
Chemistry
263,18108-18117.
FASOLATO,
C.,
PIZZO,
P.
&
POZZAN,
T.
(1990).
Receptor-mediated
calcium
influx
in
PC12
cells:
ATP
and
bradykinin
activate
two
independent
pathways.
Journal
of
Biological
Chemistry
265,
20351-20355.
FREDLHOM,
B.
B.,
ABFBRACHIO,
Ml.
P.,
BURNSTOCK,
G.,
DALY,
J.
V.,
HARDEN,
K.
T.,
JACOBSON,
K.
A.,
LEFF,
P.
&
XVILLIA-MS,
Ml.
(1994).
Nomienclature
ancd
classification
of
purinoceptors.
Pharmacological
Reviews
64,
445-475.
GRYNKIEWICZ,
C.,
POENI,
Al.
&
TsIEN,
R.
Y.
(1985).
A
new
generation
of
Ca'2
indicators
with
greatly
improved
fluorescence
properties.
Journal
of
Biological
Chemtlistry
260,
3440-3450.
HOYLE,
C.
H.
V.
&
BUERNSTOCK,
G. (1991).
ATP
receptors
and
their
physiological
roles.
In
Adenosine
in
the
Nervous
System,
ed.
STONE,
T.
W.,
pp.
43-76.
Academic
Press,
London.
JUUL,
B.,
LUSCHER,
M.
E.,
AALKJEER,
C.
&
PLESNER,
L.
(1991).
Nucleotide
hydrolytic
activity
of
isolated
intact
rat
mesenteric
small
arteries.
Biochimnica
et
Biophysica
Acta
1067,
201-207.
JUUL,
B.,
PLESNER,
L.
&
AALKJEER,
C.
(1992).
Effects
of
ATP
and
UTP
on
[Ca2+]i,
membrane
potential
and
force
in
isolated
rat
small
arteries.
Journal
of
Vascular
Research
29,
385-395.
J
Physiol.492.3
JUUL,
B.,
PLESNER,
L.
&
AALKJEER,
C.
(1993).
Effects
of
ATP
and
related
nucleotides
on
the
tone
of
isolated
rat
mesenteric
resistance
arteries.
Journal
of
Pharmacology
and
Experimental
Therapeutics
264,1234-1240.
KENNEDY,
C.
&
LEFF
P.
(1995).
How
should
P2X-purinoceptors
be
classified
pharmacologically?
Trends
in
Pharmacological
Sciences
16,
168-174.
KITAJIMA,
S.,
OZAKI,
H.
&
KARAKI,
H.
(1993).
Effects
of
ATP
and
a,f-methylene-ATP
on
cytosolic
Ca2+
level
and
force
in
rat
isolated
aorta.
British
Journal
of Pharmacology
110,
263-268.
Low,
A.
M.,
KwAN,
C.
Y.
&
DANIEL,
E.
E.
(1992).
Evidence
for
two
types
of
internal
Ca2P
stores
in
canine
mesenteric
artery
with
different
refilling
mechanisms.
A
mericani
Journal
of
Physiology
262,
H31-37.
MICKECHNIE,
K.
C.
W.,
CRACK,
B.
E.,
DAINTY,
I.
A.
&
LEFF,
P.
(1994).
The
effect
of
ecto-ATPase
inhibition
on
agonist
potency
orders
at
P2Y
and
P2X
purinoceptors.
British
Journial
of
Pharmnacology
114,
104P.
MICMIILLAN,
M.
K.,
SOLTOFF,
S.
P.,
CANTLEY,
L.
C.
&
TALAMO
B.
R.
(1987).
Extracellular
ATP
elevates
intracellular
free
calcium
in
rat
parotid
acinar
cells.
Biochemical
and
Biophysical
Research
Communications
149,
523-530.
MIERRIT,
J.
E.,
ARMSTRONG,
W.
P.,
HALLAM,
T.
J.,
JAXA-CHAMIEC,
A.,
LEIGH,
B.
K.,
i'JOORES,
K.
E.
&
RINK,
T.
J.
(1990).
SK&F
96365,
a
novel
inhibitor
of
receptor-mediated
calcium
entry.
British
Journal
of
Pharnacology
271,
515-522.
NAMIBI,
P.,
HSIAO-LING,
WV.
&
NAMBI,
A.
(1990).
Involvement
of
pertussis
toxin
sensitive
GTP
binding
protein
in
adenosine
triphosphate
and
thrombin-mediated
responses
in
vascular
smooth
muscle
cells.
In
Receptor,
ed.
LITWACK,
G.,
pp.
55-64.
Humnana
Press,
UK.
OHYA,
Y.
&
SPERELAKIS,
N.
(1989).
ATP
regulation
of
the
slowr
calcium
channels
in
v-ascular
smootlh
musele
cells
of
the
guinea-pig
mesenteric
artery.
Circulation7
Research
64,
145-154.
PACAUD,
G.,
GREGOIRE,
G.
&
LOIRAND
(1994).
Release
of
Ca
from
intracellular
store
in
smooth
muscle
cells
of
rat
poital
vein
by
ATP-
induced
Ca2+
entry.
British
Journial
of
Pharmacology
113,
457-462.
PAPP,
B.,
EYEDI,
A.,
KoVACs,
T.,
SARCADI,
S.,
WUYTACK,
F.,
THAPSTRUP,
O.,
GARDOS,
G.,
BREDOUS,
R.,
LEVY-TOLEDANO,
S.
&
ENOUF,
J.
(1991).
Demonstration
of
two
forms
of
calcium
pumps
by
thapsigargin
inhibition
and
radioimmunoblotting
in
platelet
membrane
vesicles.
Journal
of
Biological
Chemistry
266,
14593-14596.
SHIMA,
H.
&
BLAUSTEIN,
M.
(1992).
Modulation
of
evoked
contractions
in
rat
arteries
by
ryanodine,
thapsigargin
and
cyclopiazonic
acid.
Circulationi
Research
176,
117-125.
SNEDDON,
P.,
XVALKER,
P.,
LEFF,
P.
&
KENNEDY
C.
(1994).
EnhanceInent
of
syInpathetic,
purinergic
rieurotlanSmiSSion
by
a
novel
ecto-ATPase
inlhibitor
in
guinea-pig
isolated
vas
deferens.
British
Journial
of
Pharmacology
114,
109P.
SUZUKI,
H.
(1985).
Electmrical
responses
of
smnootlh
imuscle
cells
of
the
rabbit
ear
artery
to
adenosine
triphosphate.
Journial
of
Physiology
359,
401-415.
TATSURO,
Y.,
LINO,
M.,
YAMAZAWA,
T.
&
ENDO,
Al.
(1991).
Caffeine-
sensitive
and
insensitive
intracellular
calcium
stores
in
vascular
smooth
muscle
cells
of
the
guinea
pig.
Journal
of
Vascular
Medicine
and
Biology
3,154-160.
TAWADA,
Y.,
FURUKAWA,
K.
&
SHIGEKAWA,
AI.
(1987).
ATP-induced
calciumn
transient
in
cultured
aortic
smootlh
muscle
cells.
Journal
of
Biochemistry
102,
1499-1509.
G.
J
L.
Lagaud
and
others

Ca2+
and
A
TP
in
resistance
arteries
TRIBE,
R.
M.,
BoRIN,
M.
L.
&
BLAUSTEIN,
M.
P.
(1994).
Functionally
and
spatially
distinct
Ca2P
stores
are
revealed
in
cultured
vascular
smooth
muscle
cells.
Proceedings
of
the
National
Academy
of
Sciences
of
the
USA
91,
5908-5912.
VALERA,
S.,
Hussy,
N.,
EVANS,
R.
J.,
ADAMI,
N.,
NORTH,
R.
A.,
SURPRENANT,
A.
&
BUELL,
G.
(1994).
A
new
class
of
ligand-gated
ion
channel
defined
by
P2X
receptor
for
extracellular
ATP.
Nature
371,
516-519.
VON
DER
WEID,
P.
Y.,
SEREBRYAKOV,
V.
N.,
ORALLO,
F.,
BERGMANN,
C.,
SNETKOV,
V.
A.
&
TAKEDA,
K.
(1993).
Effects
of
ATP
on
cultured
smooth
muscle
cells
from
rat
aorta.
British
Journal
of
Pharmacology
108,
638-645.
VON
KtGLEGEN,
I.
&
STARKE,
K.
(1991).
Noradrenaline-ATP
co-
transmission
in
the
sympathetic
nervous
system.
Trends
in
Pharmacological
Sciences
12,
319-324.
ZIGANSHIN,
A.
U.,
HOYLE,
C.
H.
V.,
LAMBRECHT,
G.,
MUTSCHLER,
E.,
BXUMERT,
H.
G.
&
BURNSTOCK,
G.
(1994).
Selective
antagonism
by
PPADS
in
rabbit
isolated
blood
vessels.
British
Journal
of
Pharmacology
111,
923-929.
Acknowledgements
This
work
was
partially
supported
by
a
European
Union
grant
(Biomed2).
The
authors
thank
Dr
R.
Hiley
and
Dr
B.
Muller
for
fruitful
discussion
and
for
carefully
reading
the
manuscript.
Received
26
May
1995;
accepted
30
November
1995.
J
Physiol.
492.3
703
